Фотосинтез
«Биология» пәні бойынша оқу материалдары
Фотосинтез (ежелгі грек тілінен аударғанда φῶς — «жарық» және σύνθεσις — «қосылма», «бүктелу», «байланыс», «синтез») — көрінетін жарықтың (кейбір жағдайларда инфрақызыл сәулелену) энергиясын түрлендірудің күрделі химиялық процесі. фотосинтетикалық пигменттердің қатысуымен органикалық заттардың химиялық байланыстарының энергиясы (өсімдіктердегі хлорофилл, бактериялардағы бактериохлорофилл және архейлердегі бактериородопсин).
Фотосинтездің мәні
Фотосинтез, химосинтетикалық бактерияларды қоспағанда, планетадағы барлық тіршіліктің энергетикалық негізін құрайды.
Фотосинтез құрлықтағы өсімдіктердің жасыл бөліктерінде және балдырларда жүреді. Бір жылда жасыл балдырлар Жер атмосферасына 3,6⋅10**11 тонна оттегі бөледі, бұл жердегі фотосинтез кезінде түзілетін барлық оттегінің 90% құрайды. Фотосинтез — жер бетіндегі ең кең таралған биохимиялық процесс.
Жер бетінде 3 миллиард жылдан астам уақыт бұрын су молекуласының күн сәулесінің кванттары бойынша О2 түзілуімен бөліну механизмінің пайда болуы биологиялық эволюцияның ең маңызды оқиғасы болып табылады, ол Күн сәулесін энергияның негізгі көзіне айналдырды. биосферада.
Фототрофтар Күнде болатын термоядролық реакциялардың энергиясының органикалық молекулалардың энергиясына айналуын және сақталуын қамтамасыз етеді. Күн энергиясы фототрофтардың қатысуымен органикалық заттардың химиялық байланыстарының энергиясына айналады. Гетеротрофты организмдердің болуы тек фототрофтардың органикалық қосылыстарда жинақталған энергиясының арқасында мүмкін болады. Органикалық заттардың химиялық байланыстарының энергиясын пайдаланған кезде гетеротрофтар оны тыныс алу және ашыту процестерінде босатады.
Фотосинтез ауылшаруашылық өсімдіктерінің де, жануарлардың тағамдық өнімдерінің де өнімділігінің негізі болып табылады.
Адамзаттың биоотын (ағаш, түйіршіктер, биогаз, биодизель, этанол, метанол) және қазба отындарын (көмір, мұнай, табиғи газ, шымтезек) жағу арқылы алған энергиясы да фотосинтез процесінде жинақталады.
Фотосинтез биогеохимиялық циклге бейорганикалық көміртектің негізгі кірісі ретінде қызмет етеді.
Атмосферадағы бос оттегінің көп бөлігі биогенді болып табылады және фотосинтездің жанама өнімі болып табылады. Тотықтырғыш атмосфераның пайда болуы (оттегі апаты) жер бетінің күйін толығымен өзгертті, тыныс алудың пайда болуын мүмкін етті, ал кейінірек озон қабаты пайда болғаннан кейін құрлықта өмір сүруге мүмкіндік берді.
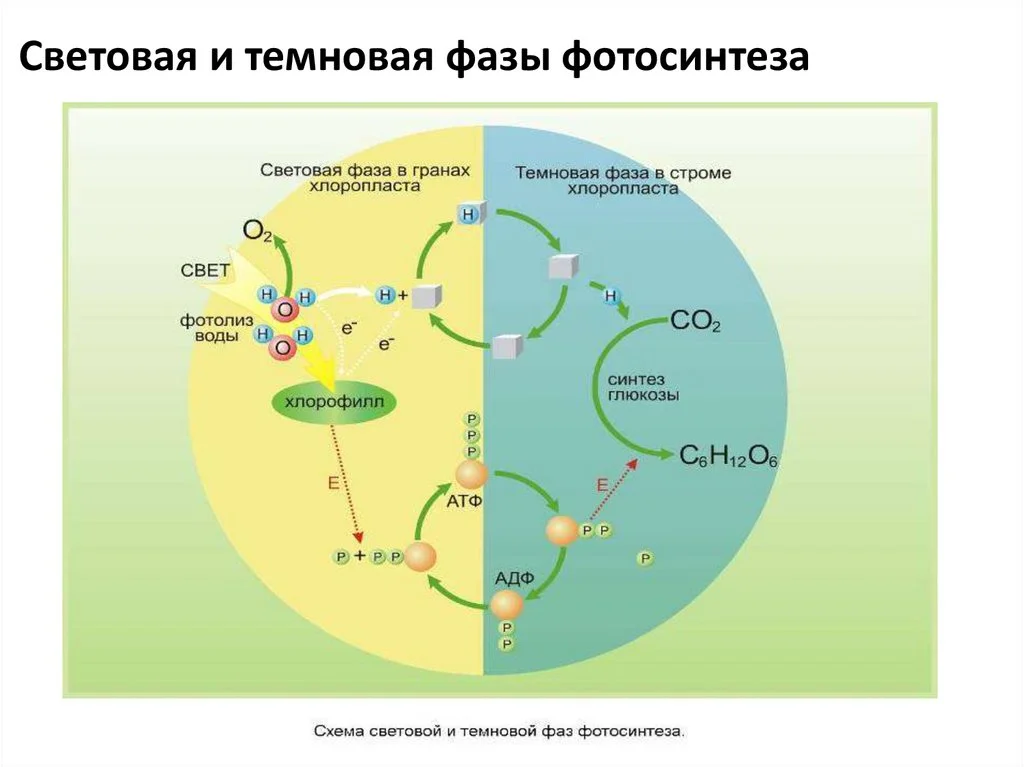
Өсімдіктерде жүретін фотосинтез процесінің схемалық бейнесі
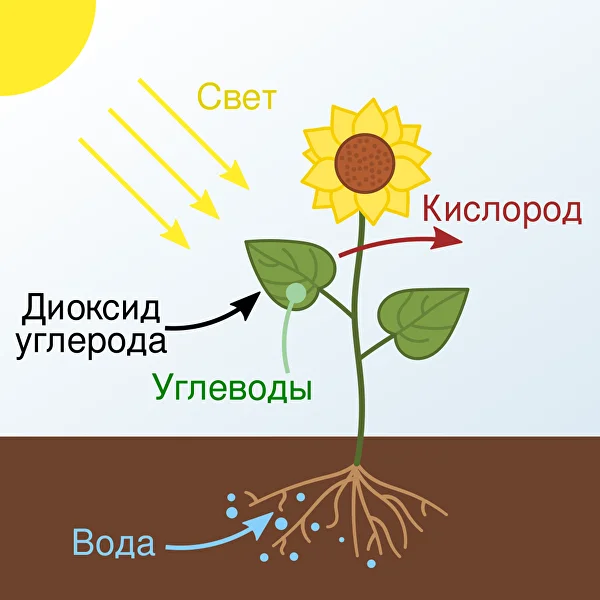
Фотосинтез барлық өсімдіктер үшін органикалық заттардың негізгі көзі, сонымен қатар көптеген жануарлардың қоректенуінің негізі болып табылады. Фотосинтез деп жарықтың әсерінен жасыл өсімдіктер мен кейбір бактериялардың жасушаларында болатын химиялық реакцияларды айтады.
Фотосинтез кезінде жарық энергиясы химиялық энергияға айналады, ол глюкоза деп аталатын органикалық молекулаларды синтездеуге жұмсалады. Бұл процестің негізгі құрамдас бөлігі әртүрлі толқын ұзындығындағы жарықты жұтуға қабілетті хлорофилл пигменттері болып табылады. Хлорофилл жарықты жұтқанда, фотожүйелердің электрондық тізбектері бойымен қозғала бастайтын электрондарды шығарады. Бұл су молекулаларының оттегі мен протондарға ыдырауына, сондай-ақ жасушаларда энергияның негізгі тасымалдаушысы АТФ түзілуіне әкеледі.
Фотосинтез тек өсімдіктер үшін маңызды процесс емес, сонымен қатар бүкіл биосфера үшін ғаламдық маңызы бар. Ол жердегі тіршілікке қажетті оттегінің түзілуін қамтамасыз етеді, сонымен қатар атмосферадағы көмірқышқыл газының концентрациясын азайтады. Сонымен қатар, фотосинтез барлық ағзаларды энергиямен қоректендіретін қоректік тізбекте негізгі рөл атқарады. Фотосинтез механизмін зерттеу тірі жүйелердің қалай жұмыс істейтінін түсінуге көмектеседі және күн энергиясын пайдаланудың жаңа әдістерін жасау үшін үлкен маңызға ие болуы мүмкін.
Фотосинтез үшін организмдерге арнайы пигменттер мен фотосинтетикалық қабылдағыштар (антенналар) қажет. Антенналар жарық кванттарын жұтып, оларды энергияға айналдырады, содан кейін ол дененің мембраналарында H+ иондарының электрохимиялық градиентін жасау үшін пайдаланылады.
Өсімдіктер мен балдырлар сияқты тірі ағзалардың көпшілігінде хлорофиллдер антенна рөлін атқарады. Хлорофиллдер — көрінетін диапазондағы жарықты сіңіретін және оның энергиясын фотосинтезге беретін негізгі пигменттер. Дегенмен, антенна ретінде А дәруменінің туындысын, ретиналды қолданатын басқа да организмдер бар. Бұл сирек кездесетін жағдай, бірақ ол әлі де бар.
Сонымен, фотосинтездің екі түрін ажыратуға болады: хлорофилл және хлорофилл емес. Хлорофилл фотосинтезінде хлорофиллдер жарықты жұтып, оны энергияға айналдыруда шешуші рөл атқарады. Хлорофиллсіз фотосинтезде сетчатка негізіндегі антенналар ұқсас функцияны орындайды.
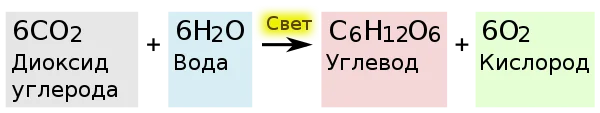
Өсімдіктерде жүретін фотосинтездің жеңілдетілген теңдеуі
Жарық (жарыққа тәуелді) кезең
Фотосинтездің жеңіл сатысында жоғары энергиялы өнімдер түзіледі: жасушада энергия көзі қызметін атқаратын АТФ және тотықсыздандырғыш ретінде қолданылатын НАДФ. Жанама өнім ретінде оттегі бөлінеді.
Процестің фотохимиялық мәні
Хлорофиллдің қозуының екі деңгейі бар (бұл оның жұтылу спектрінде екі максимумның болуына байланысты): біріншісі конъюгацияланған қос байланыстар жүйесінің электронының жоғары энергетикалық деңгейіне өтуімен байланысты, екіншісі порфирин ядросының азот пен магнийдің жұпталмаған электрондарының қозуы. Электронның спині өзгеріссіз қалғанда бірінші және екінші синглетті қозған күйлер, ал электрон спинін өзгерткенде бірінші және екінші триплет күйлері пайда болады.
Екінші қозған күй ең жоғары энергиялы, тұрақсыз және хлорофилл одан біріншіге 10−12 с ішінде тек жылу түрінде 100 кДж/моль энергия жоғалтумен ауысады. Бірінші синглет және триплет күйлерінен молекула жарық түріндегі энергияның бөлінуімен (сәйкесінше флуоресценция және фосфоресценция) немесе энергияның басқа молекулаға ауысуымен жылумен немесе электрон болғандықтан негізгі күйге ауыса алады. жоғары энергия деңгейі электронның басқа қосылысқа ауысуымен ядромен әлсіз байланысқан.
Бірінші мүмкіндік жарық жинайтын кешендерде, екіншісі — реакция орталықтарында жүзеге асады, мұнда жарық кванты әсерінен қозған күйге өтетін хлорофилл электрон донорына (тотықсыздандырғыш) айналады және оны бастапқы акцепторлар. Электронның оң зарядталған хлорофиллге оралуына жол бермеу үшін біріншілік акцептор оны екіншіге ауыстырады. Сонымен қатар, алынған қосылыстардың өмір сүру ұзақтығы қозғалған хлорофилл молекуласына қарағанда жоғары. Энергияны тұрақтандыру және зарядты бөлу орын алады. Одан әрі тұрақтандыру үшін екіншілік электрон доноры оң зарядталған хлорофиллді азайтады, ал оттегі фотосинтезі кезіндегі біріншілік донор су болып табылады.
Оттегі фотосинтезін жүргізетін организмдердің алдында тұрған мәселе судың тотығу-тотықсыздану потенциалдарының айырмашылығы (H2O → O2 жартылай реакциясы үшін (E0 = +0,82 В) және NADP+ (E0 = -0,32 В). Бұл жағдайда хлорофиллдің болуы керек. суды тотықтыру үшін негізгі күйде потенциалы +0,82 В-тан жоғары, бірақ сонымен бірге NADP+ азайту үшін қозған күйде потенциалы бар, сондықтан екі талапқа жауап бере алмайды фотожүйелер пайда болды және процесті аяқтау үшін әртүрлі типтегі екі жарық кванттары және екі хлорофилл қажет.
Жеңіл жинау кешендері
Хлорофилл екі қызмет атқарады: энергияны сіңіру және беру. Хлоропластардың жалпы хлорофиллінің 90%-дан астамы I немесе II фотожүйелердің реакциялық орталығына энергияны жіберетін антенна қызметін атқаратын жарық жинау кешендерінің (LHC) бөлігі болып табылады. Хлорофиллден басқа, SSC құрамында каротиноидтар бар, ал кейбір балдырлар мен цианобактерияларда фикобилиндер бар, олардың рөлі хлорофилл салыстырмалы түрде әлсіз сіңіретін толқын ұзындығының сәулесін жұту болып табылады.
Энергияның берілуі резонансты жол (Фёрстер механизмі) арқылы жүреді және бір жұп молекула үшін 10−10–10−12 с уақытты алады, оның өтуі шамамен 1 нм болады; Тасымалдау кейбір энергия жоғалтуларымен (10% хлорофилл а-дан хлорофиллге, 60% каротиноидтардан хлорофиллге) жүреді, сондықтан қысқа толқын ұзындығында максималды сіңіру қабілеті бар пигменттен ұзағырақ пигментке ғана мүмкін болады. . Дәл осы тәртіпте SSC пигменттері өзара локализацияланады, ең ұзын толқын ұзындығы хлорофиллдер реакция орталықтарында орналасады. Кері энергияның ауысуы мүмкін емес.
Өсімдіктердің СКҚ цианобактериялардағы тилакоидтардың қабықшаларында орналасады, оның негізгі бөлігі мембраналардан тыс оларға бекінген фикобилисомаларға – таяқша тәрізді полипептидті-пигменттік кешендерге тасымалданады, оларда әртүрлі фикобилиндер орналасады: шетінде фикоэритриндер бар; (абсорбциялық максимум 495-565 нм-де), олардың артында реакция орталығының хлорофилл а (680-700 нм) энергиясын тізбектей беретін фикоцианиндер (550-615 нм) және аллофикоцианиндер (610-670 нм) тұрады.
Электронды тасымалдау тізбегінің негізгі компоненттері
Фотожүйе II
Фотожүйе — бұл СКҚ, фотохимиялық реакция орталығы және электронды тасымалдаушылардың жиынтығы. Жарық жинау кешені II құрамында 200 молекула хлорофилл а, 100 молекула хлорофилл b, 50 молекула каротиноид және 2 молекула феофитин бар. II фотожүйенің реакция орталығы тилакоидты мембраналарда орналасқан және СКҚ қоршалған пигменттік-белоктық кешен. Оның құрамында 680 нм (P680) максималды сіңіру бар хлорофилл-а димер бар. Жарық квантының СКҚ энергиясы ақырында оған ауысады, нәтижесінде электрондардың біреуі жоғары энергетикалық күйге ауысады, оның ядромен байланысы әлсірейді, ал қозғалған Р680 молекуласы күшті тотықсыздандырғышқа айналады ( химиялық потенциал E0 = −0,7 В).
P680 феофитинді төмендетеді, содан кейін электрон PS II құрамына кіретін хинондарға, содан кейін b6f кешеніне қалпына келтірілген түрде тасымалданатын пластокинондарға ауысады. Пластокинонның бір молекуласында стромадан алынған 2 электрон және 2 протон болады.
Р680 молекуласындағы электронды бос орынның толтырылуы судың есебінен болады. PS II белсенді орталығында 4 марганец ионынан тұратын су тотықтырғыш кешенін қамтиды. Бір оттегі молекуласын құру үшін 4 электрон беретін екі су молекуласы қажет. Сондықтан процесс 4 циклде жүзеге асырылады және оның толық орындалуы үшін 4 квант жарық қажет. Кешен интратилакоидты кеңістіктің бүйірінде орналасады және оған пайда болған 4 протон бөлінеді.
Сонымен, PS II жұмысының жалпы нәтижесі 4 жарық квантының көмегімен интратилакоидты кеңістікте 4 протонның және мембранада 2 қалпына келтірілген пластокинонның түзілуімен 2 су молекуласының тотығуы болып табылады.
b6/f цитохромы кешені
b6f кешені протондарды стромадан интратилакоидтық кеңістікке айдайтын және электронды тасымалдау тізбегінің тотығу-тотықсыздану реакцияларында бөлінетін энергия есебінен олардың концентрациясының градиентін жасайтын сорғы болып табылады. 2 пластокинон 4 протонды айдайды. Кейіннен трансмембраналық протондық градиент (строманың рН шамамен 8, интратилакоидтық кеңістіктің рН 5) трансмембраналық фермент АТФ синтаза арқылы АТФ синтезі үшін қолданылады.
Фотожүйе I
Жарық жинау кешені I шамамен 200 хлорофилл молекуласын қамтиды.
Бірінші фотожүйенің реакция орталығында абсорбциялық максимумы 700 нм (Р700) болатын хлорофилл а димері бар. Жарық квантымен қозудан кейін ол екінші реттік акцепторды қалпына келтіретін хлорофилл а-ны қалпына келтіреді (К1 витамині немесе филлохинон), содан кейін электрон ферредоксин-НАДФ редуктаза ферментінің көмегімен НАДФ-ны төмендететін ферредоксинге ауысады.
b6f кешенінде тотықсызданған пластоцианин ақуызы интратилакоидты кеңістік жағынан бірінші фотожүйенің реакциялық орталығына тасымалданады және тотыққан P700-ге электрон береді.
Циклдік және псевдоциклдік электронды тасымалдау
Жоғарыда сипатталған толық циклдік емес электрон жолынан басқа, циклдік және псевдоциклдік жол ашылды.
Циклдік жолдың мәні ферредоксин НАДФ орнына пластохинонды төмендетеді, ол оны қайтадан b6f кешеніне ауыстырады. Бұл үлкен протон градиентіне және көбірек ATP-ге әкеледі, бірақ NADPH жоқ.
Псевдоциклдік жолда ферредоксин оттегін төмендетеді, ол одан әрі суға айналады және II фотожүйеде қолданылуы мүмкін. Бұл жағдайда NADPH да түзілмейді.
Қараңғы фаза
Қараңғы кезеңде АТФ және НАДФ қатысуымен СО2 глюкозаға дейін тотықсызданады (C6H12O6). Бұл процесс үшін жарық қажет болмаса да, оны реттеуге қатысады.
С3 фотосинтезі, Кальвин циклі
Кальвин циклі немесе редукциялық пентозофосфат циклі үш кезеңнен тұрады:
- карбоксилдену;
- қалпына келтіру;
- CO2 акцепторының регенерациясы.
Бірінші қадамда СО2 рибулоза-1,5-бисфосфатқа рибулоза-бисфосфат карбоксилаза/оксигеназа ферменті арқылы қосылады. Бұл ақуыз хлоропласт ақуыздарының негізгі бөлігін құрайды және табиғатта ең көп таралған фермент болып саналады. Нәтижесінде 3-фосфоглицерин қышқылының (ПГА) екі молекуласына ыдырайтын тұрақсыз аралық қосылыс түзіледі.
Екінші кезеңде FHA екі кезеңде қалпына келтіріледі. Біріншіден, ол 1,3-бисфосфоглицерин қышқылының (ДПГА) түзілуімен фосфороглицерокиназаның әсерінен АТФ арқылы фосфорланады, содан кейін триосфосфатдегидрогеназа және НАДФН әсерінен ДПГА ацилфосфат тобы фосфорсызданады және тотықсызданады. альдегид және глицеральдегид-3-фосфат — фосфорланған көмірсулар (ФКА) түзіледі.
Үшінші кезеңде 5 PHA молекуласы қатысады, олар 4-, 5-, 6- және 7-көміртекті қосылыстардың түзілуі арқылы 3 5-көміртекті рибулоза-1,5-бифосфатқа біріктіріледі, ол үшін 3 АТФ қажет.
Ақырында, глюкоза синтезі үшін екі PHA қажет. Оның молекулаларының бірін қалыптастыру үшін циклдің 6 айналымы, 6 СО2, 12 NADPH және 18 АТФ қажет.
C4 фотосинтезі
Бұл фотосинтез механизмінің әдеттегіден айырмашылығы көмірқышқыл газын бекіту және оны пайдалану өсімдіктің әртүрлі жасушалары арасындағы кеңістікте бөлінеді.
Стромада еріген СО2 төмен концентрациясында рибулоза-бифосфаткарбоксилаза рибулоза-1,5-бифосфаттың тотығу реакциясын катализдейді және оның 3-фосфоглицерин қышқылына және фосфогликоль қышқылына ыдырайды, оны фототыныс алу процесінде қолдануға мәжбүр етеді.
CO2 концентрациясын арттыру үшін C4 өсімдіктері жапырақ анатомиясын өзгертті. Кальвин циклі қантамырлар шоғырының қабықша жасушаларында локализацияланады, ПЭП карбоксилазасының әсерінен фосфоэнолпируват карбоксилденеді, ол малатқа немесе аспартатқа айналады және қабықша жасушаларына тасымалданады; декарбоксилденген пируват түзеді, ол мезофилл жасушаларына қайтарылады.
С4 фотосинтезі іс жүзінде Кальвин циклінен рибулоза-1,5-бифосфаттың жоғалуымен қатар жүрмейді, сондықтан тиімдірек. Бірақ 1 глюкоза молекуласының синтезі үшін 18 емес, 30 АТФ қажет. Бұл тропиктік аймақтарда ақталған, онда ыстық климат устьицаларды жабық ұстауды талап етеді, бұл СО2-нің жапыраққа енуіне жол бермейді, сондай-ақ өмірдің өрескел стратегиясымен.
7600-ге жуық өсімдік түрі фотосинтезді С4 жолы арқылы жүзеге асырады. Олардың барлығы гүлділер тұқымдасына жатады: көптеген дәнді дақылдар (61% түрлер, оның ішінде мәдени – жүгері, қант қамысы, құмай және т.б.), гвоздика (Chenopoaceae тұқымдасында ең көп үлес – 40% түр, Amaranthaceae – 25). %), кейбір қияқтар, астрагүлділер, брассикалар, эйфорбиялар.
CAM фотосинтезі
CAM типті фотосинтезде (Crassulaceae қышқылының метаболизмі) СО2 ассимиляциясының бөлінуі және Кальвин циклі С4 сияқты кеңістікте емес, уақыт бойынша жүреді. Түнде малат жасушалардың вакуольдерінде жинақталады, жоғарыда сипатталғанға ұқсас механизм бойынша, күндіз ашық устьицалармен, Кальвин циклі жүреді; Бұл механизм суды мүмкіндігінше үнемдеуге мүмкіндік береді, бірақ тиімділігі жағынан C4 және C3 екеуінен де төмен. Бұл стресске төзімді өмірлік стратегиямен негізделген.